theorists, while not accepting our preference for viewing stasis in the
context or habitat tracking 17 or developmental constraint 33,34,
have been persuaded by punctuated equilibrium that maintenance of stability
within species must be considered as a major evolutionary problem.
Macroevolution as a problem in species sorting. If punctuated equilibrium
has provoked a shift in paradigms for macroevolutionary theory (see ref.
35 for a defence of this view), the main insight for revision holds that
all substantial evolutionary change must be reconceived as higher-level
sorting based on differential success of certain kinds of stable species,
rather than as progressive transformation within lineages (see Eldredge
36 on taxic versus transformational views of evolution; Simpson
37, however, in the canonical paleontological statement of the
generation before punctuated equilibrium, had attributed 90% of macroevolution
to the transformational mode, and only 10%. to speciation). Figure 1, our
original diagram of punctuated equilibrium, shows how a trend may be produced
by differential success of certain species without directional change in
any species following its origin.
Darwin's theory of natural selection locates the causality of evolutionary
change at one domain on one level: natural selection operating by struggle
among individual organisms for reproductive success. Given Darwin's crucial
reliance upon lyellian uniformity for extrapolating this mode of change
to encompass all magnitudes through all times, the interposition of a
level for sorting among stable species breaks this causal reduction and
truly, in Stanley's felicitous term 38, "decouples"
macro- from microevolution. Decoupling is not a claim for the falseness
or irrelevancy of microevolutionary mechanisms, especially natural selection,
but a recognition that Darwinian extrapolation cannot fully explain large-scale
change in the history of life.
The main point may be summarized as follows. Most macro-evolution must
be rendered by asking what kinds of species within a clade did better
than others (speciated more frequently, survived longer), or what biases
in direction of speciation prevailed among species within a clade. Such
questions enjoin a very different programme of research from the traditional
how did natural selection within a lineage build substantial adaptation
during long stretches of time?' The new questions require a direct study
of species and their differential success, older queries focused downward
upon processes within populations and their extrapolation through time.
Darwin's location of causality in organisms must be superseded by a hierarchical
model of selection, with simultaneous and important action at genie, organismal
and taxic levels 39,40. Williams 34, who so stoutly
defended classical Darwinism against older, invalid, and very different
forms of group selection 41, now acknowledges the importance
of such clade selection in macroevolution. Punctuated equilibrium has
been used as a central concept in the development of hierarchy theory
in evolutionary biology.
Implications. Any theory with a claim to novelty in broad perspective
must enlighten old problems and suggest extensions. The speciational view
of macroevolution, which does not strictly require punctuated equilibrium,
but which was nurtured and has thrived in its context, requires a reformulation
of nearly all macroevolutionary questions. For example, so-called living
fossils, once treated as lineages rendered static by optimal adaptation,
unusually Stable environment, or lack of genetic variation, should be reconceptualized
as members of groups with unusually low speciation rates, and therefore
little opportunity to accumulate change 42, (We have no evidence
that the species of 'living fossil' groups are particularly old. For example,
the western Atlantic horseshoe crab, Limulus polyphemus--the
'type example' of the phenomenon--has no fossil record at all, whereas
the genus can only be traced to the Miocene.)
Going further, the entire tradition of expressing evolutionary change
in darwin units (where 1 darwin equals character change by a factor of
e in 1 million years) 43 makes no sense in a speciational context.
(If a lineage goes from species A to D in 10 million years through three
episodes of rapid change with intervening stasis, a cited rate of so many
millidarwins becomes a meaningless average,) We learn as a received truth
of evolution, for example, that human brain size increased at an extra-ordinary
(many say unprecedented) rate during later stages of our lineage. But
this entrenched belief may be a chimaera born of an error in averaging
rates over both punctuations and subsequent periods of stasis. Homo
sapiens is a young species, perhaps no more than 200,000 years old.
If most of our increment accrued quickly at our origin, but we then express
this entirety from our origin to the present time as a darwin rate, we
calculate a high value because our subsequent time of stasis has been
so short. But if the same speciation event, with the same increment in
the same time, had occurred two million years ago (with subsequent stasis),
the darwin rate for the identical event would be much lower.
Cope's rule, the tendency for phyletic increase in body size, had generally
been attributed to selective value of large size within anagenetic lineages,
but is probably better interpreted 44,45 as greater propensity
for speciation in smaller species, for whom increasing size is the only
'open' pathway (see Martin 46 on the negative correlation of
generic species richness and body size). Raup and Sepkoski 47
proposed a conventional explanation for decreasing rate of background
extinction through geological time: generally better adaptation of later
species. But Valentine 48 and Gilinsky (personal communication)
offer an interesting speciational alternative: if extinction intensities
were constant through time, groups with equally high speciation and extinction
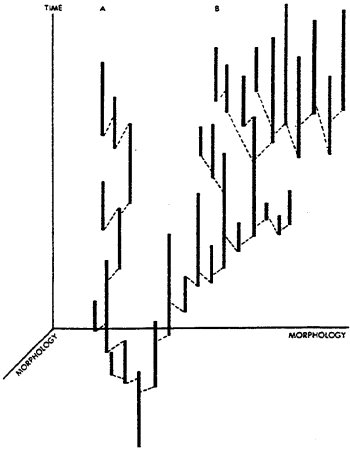
FIG. 1 Three-dimensional sketch contrasting a pattern of relative stability
(A) with a trend (B), where speciation (dashed lines) is occurring in
both major lineages. Morphological change is depicted here along the horizontal
axes, while the vertical axis is time. Though a retrospective pattern
of directional selection might be fitted as a straight line in (B), the
actual pattern is stasis within species, and differential success of species
exhibiting morphological change in a particular direction. For further
explanation, see ref. 1.
| 224 |
NATURE - VOL 366 - 18 NOVEMBER 1993 |
|